Rastlinné organizmy sú komplexné systémy, ktoré neustále vykonávajú nespočetné množstvo fyziologických a biochemických procesov nevyhnutných pre ich život. Tieto procesy možno rozdeliť do dvoch hlavných kategórií: primárny metabolizmus a sekundárny metabolizmus.
Úvod do rastlinného metabolizmu
Zatiaľ čo primárny metabolizmus zahŕňa základné biochemické dráhy nevyhnutné pre rast, vývoj a reprodukciu rastlín, ako je fotosyntéza a dýchanie, sekundárny metabolizmus sa zameriava na produkciu širokej škály špecializovaných organických zlúčenín, ktoré nie sú priamo zapojené do týchto základných procesov.
Primárny metabolizmus rastlín
Primárny metabolizmus zahŕňa procesy, ktoré sú esenciálne pre prežitie a rast rastliny. Ide o reakcie, ktoré sú univerzálne pre väčšinu organizmov a sú nevyhnutné pre produkciu energie a stavebných blokov buniek.
Fotosyntéza - Kľúčový primárny proces
Pojem autotrofia označuje schopnosť organizmu vyrobiť si vlastné organické látky z jednoduchých anorganických zlúčenín (CO₂ a H₂O). Autotrofné organizmy (predovšetkým rastliny) sú tak primárnymi producentmi biosféry, od ktorých priamo či nepriamo závisí zvyšok planéty.
Fotosyntéza je fundamentálny dej, pri ktorom sa svetelná energia Slnka premieňa na stabilnú energiu chemických väzieb organických molekúl. Vedľajším produktom tohto procesu je kyslík.
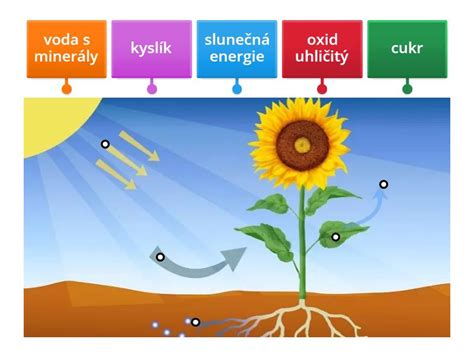
Primárne procesy fotosyntézy (Svetelná fáza)
Primárne procesy fotosyntézy vyžadujú prítomnosť svetla, preto bývajú označované ako fotochemická fáza. Základnou úlohou primárnych procesov je premeniť kinetickú energiu fotónov (svetla) na využiteľnú chemickú energiu vo forme molekúl ATP a redukčného činidla NADPH + H⁺. U fotosyntetizujúcich rastlín prebiehajú tieto deje v chloroplastoch na membránach tylakoidov.
Hlavným asimilačným farbivom je chlorofyl a, ktorý sa nazýva aj aktívny chlorofyl. Ten prostredníctvom fotooxidácie (uvoľnením elektrónu) premieňa svetlo na chemickú energiu. Ostatné pigmenty (chlorofyl b, karotenoidy) fungujú ako pomocné svetlozberné antény. Okrem chlorofylu sa fotosyntézy zúčastňujú aj iné farbivá ako napríklad karotenoidy a u siníc, červených rias a kryptomonád aj fykobilíny - fykoerytríny a fykocyaníny.
Necyklický transport elektrónov prebieha za účasti oboch fotosystémov. Najprv za účasti PSII dochádza k excitácii chlorofylu v reakčnom centre P680. Zároveň dochádza k aktivácii enzýmu, ktorý rozkladá vodu na protóny, elektróny a kyslík, ktorý uniká do atmosféry. Keďže tento rozpad vody prebieha za prítomnosti svetla, označuje sa termínom fotolýza vody (Hillova reakcia):
\( \ce{2H2O -> O2 + 4H+ + 4e-} \)
Elektróny z fotolýzy vody dopĺňajú chýbajúce miesto v excitovanom P680. Vzbudené elektróny z P680 sa prenášajú ďalej cez cytochróm b6/f na reakčné centrum PSI. Energia, ktorú pri tom elektrón vyžiari, slúži na vznik protónového gradientu, ktorý následne enzým ATP syntáza využíva na syntézu ATP. Tomuto procesu hovoríme necyklická fotofosforylácia.
Cyklický transport elektrónov prebieha len za účasti PSI. Začína pohltením svetelnej energie molekulou chlorofylu v reakčnom centre P700. Z chlorofylu sa tým uvoľnia elektróny, ktoré zachytí oxidačno-redukčný enzým feredoxín. Z neho sa elektróny prenášajú späť na chlorofyl cez plastocyanín a cytochrómový komplex b₆/f. Keďže pri tomto procese vykonávajú elektróny cyklus (chlorofyl - feredoxín - oxidačno-redukčné enzýmy - chlorofyl), označujeme tento dej ako cyklická fotofosforylácia. Cyklický transport elektrónov predstavuje alternatívnu dráhu, ktorú chloroplast volí v prípade nedostatku NADP a ATP.
Presne rovnakým spôsobom vykonávajú fotosyntézu aj sinice, ktoré sú prokaryotické organizmy. Zvyšné skupiny fotoautotrofných baktérií (napr. purpurové alebo zelené sírne baktérie) už vykonávajú bezkyslíkatú (anoxygénnu) fotosyntézu. Ako zdroj vodíka na redukciu CO₂ nevyužívajú vodu, ale iné zlúčeniny, najčastejšie sírovodík (H₂S), preto pri ich fotosyntéze nevzniká kyslík, ale uvoľňuje sa síra. Mnohé purpurové baktérie taktiež redukujú NAD⁺ na NADH + H⁺, na rozdiel od rastlín a siníc, ktoré redukujú NADP⁺ na NADPH.
Sekundárne procesy fotosyntézy (Tmavá fáza - Calvinov cyklus)
Sekundárne procesy priamo nevyžadujú prítomnosť svetla, a sú preto označované ako tmavá alebo termochemická fáza fotosyntézy. Počas týchto procesov dochádza k fixácii CO₂ a vzniku sacharidov. Zdrojom energie na túto premenu je ATP a redukovadlom NADPH + H⁺.
Pomenovanie tmavá fáza nie je úplne šťastné, pretože to evokuje, že tento dej prebieha v tme. Nie je to však pravda. Hoci je tento dej nezávislý od priameho slnečného žiarenia, vyžadované sú pri ňom východiskové produkty z primárnych procesov fotosyntézy − ATP a NADPH. Cesta fixácie CO₂ týchto rastlín sa nazýva Calvinov cyklus. Reakcie možno rozdeliť do 3 fáz:
- Karboxylačná fáza − primárnym akceptorom CO₂ je ribulóza-1,5-bisfosfát (RuBP), čím vzniká nestabilná 6-uhlíkatá molekula, ihneď sa hydrolyzujúca na 2 3-uhlíkaté (preto C3) molekuly kyseliny 3-fosfoglycerovej. Reakciu väzby CO₂ na RuBP katalyzuje enzým rubisco (ribulóza-1,5-bisfosfát karboxyláza/oxygenáza), ktorý v listoch tvorí 30-50 % všetkých prítomných bielkovín. Katalytická aktivita rubisco je podmienená prítomnosťou Mg²⁺.
- Redukčná fáza − kyselina 3-fosfoglycerová sa redukuje na sacharid.
- Regeneračná fáza − regenerácia RuBP, aby cyklus mohol pokračovať.
Fotorespirácia
Fotorespirácia je proces, kedy rastlina spotrebováva O₂ a produkuje CO₂. Je to teda opačný proces k fotosyntéze. Od dýchania sa odlišuje tým, že pri fotorespirácii sa energia v podobe ATP a redukovadlo NADPH spotrebováva. Prvotnú reakciu fotorespiračnej kaskády paradoxne zahajuje taktiež enzým rubisco, avšak ako substrát používa O₂ namiesto CO₂ (preto sa nazýva karboxyláza (viaže CO₂)/oxygenáza (viaže O₂)).
V oxygenázovej reakcii reaguje O₂ s RuBP a vzniká kyselina 3-fosfoglycerová a 2-fosfoglykolát, ktorý sa ďalej enzymaticky metabolizuje až na CO₂. Tieto deje prebiehajú čiastočne aj v peroxizómoch a mitochondriách za spotreby ATP a NADPH. Úloha fotorespirácie nie je uspokojivo vysvetlená, ale predpokladá sa, že by mohla slúžiť k ochrane rastlín pred nežiadúcimi účinkami nadmerného slnečného žiarenia pri súčasnej absencii CO₂. Pri fotorespirácii zároveň vznikajú aj prekurzory pre syntézu niektorých aminokyselín (glycín, serín). Možná je aj hypotéza, že afinita O₂ k enzýmu rubisco vznikla ešte v časoch, kedy atmosféra obsahovala veľmi málo O₂, a teda neexistovala negatívna selekcia proti tomuto fenoménu.
Adaptácie fotosyntézy: C4 a CAM rastliny
Anatómia listov týchto rastlín sa odlišuje od C3 rastlín a dochádza aj k priestorovému oddeleniu fixácie CO₂ (bunky mezofylu) a jeho redukcie na glukózu (bunky pošiev cievnych zväzkov). Takto si pre enzým Rubisco udržiavajú umelo obrovskú lokálnu koncentráciu CO₂, čím úplne zabránia fotorespirácii a zachovajú si masívne výnosy cukrov.
Primárnym akceptorom CO₂ v mezofyle listov je fosfoenolpyruvát (PEP), pričom jeho karboxyláciou vzniká 4-uhlíkatá (preto C4) kyselina oxaloctová, ktorá sa ďalej pomocou NADPH + H⁺ redukuje na malát. Malát sa transportuje do pošiev cievnych zväzkov, kde sa dekarboxyluje a vzniknutý CO₂ vstupuje do Calvinoho cyklu.
Sukulenty a kaktusy z extrémnych púští šetria vodou tak, že okrem priestorového oddelenia fixácie a redukcie CO₂ oddelili procesy aj časovo. Tieto tzv. CAM rastliny (angl. Crassulaceae Acid Metabolism) predstavujú variant C4 rastlín, ktoré majú cez deň zatvorené prieduchy, aby predišli stratám vody. Zároveň však neprijímajú ani CO₂.
CAM rastliny teda prijímajú CO₂ v noci a v cytoplazme ho pomocou PEP-karboxylázy viažu na fosfoenolpyruvát (PEP). Vzniknutá kyselina oxaloctová sa pomocou NADPH + H⁺ redukuje na malát. PEP a NADPH + H⁺ sa tvoria procesom glykolýzy, takže súčasne sa znižuje obsah škrobu v chloroplastoch. Malát sa transportuje do vakuoly a okysľuje bunkovú šťavu.
Kľúčové faktory ovplyvňujúce fotosyntézu
- Svetlo - primárny zdroj energie. Najdôležitejšie je svetlo v červenej a modrofialovej časti spektra (okolo 680 a 700 nm). Rastlina dokáže zo svetla, čo na ňu dopadne využiť asi 2 %. Ostatné svetlo sa odráža, alebo prepúšťa. Pri priveľmi silnom žiarení sa fotosyntéza zastavuje kvôli poškodeniu buniek reaktívnym kyslíkom.
- Oxid uhličitý (CO₂) - kľúčový zdroj uhlíka. Z jedného gramu CO₂ sa vytvorí asi 0,5 g sušiny. Bežné rastliny dokážu zvyšovať efektivitu fotosyntézy priemerne až do koncentrácie 0,05 % CO₂ vo vzduchu.
- Voda (H₂O) - priamy zdroj elektrónov a protónov pri fotolýze. Jej nedostatok spôsobí zatvorenie prieduchov a zablokuje prísun CO₂, čím sa fotosyntéza spomalí. CAM rastliny majú niekoľkonásobne nižší prírastok biomasy práve z tohto dôvodu.
- Teplota prostredia - reguluje aktivitu enzýmov. U väčšiny našich rastlín prebieha fotosyntéza v rozmedzí 0−40 °C, no optimum je zvyčajne 25-30 °C.
Mixotrofia ako špecializovaný metabolizmus
Mixotrofia je špecializovaný spôsob výživy spájajúci autotrofiu a heterotrofiu. Rastliny normálne fotosyntetizujú, no popritom aktívne prijímajú iné organické živiny z tiel živočíchov. Tento jav je typický pre mäsožravé rastliny. Žijú v drsných a podmáčaných biotopoch (rašeliniská), kde je v pôde pre absenciu užitočných baktérií kritický nedostatok dusíka. Deficit dusíka pre tvorbu bielkovín si preto kompenzujú lovom hmyzu pomocou premenených listov.
Jednotlivé druhy si vyvinuli rôzne typy pascí:
- Bublinatka (Utricularia) loví vo vode pomocou malých komôrok, ktoré hmyz pri podráždení bleskovo nasajú podtlakom.
- Rosička (Drosera) má na listoch lepivé trichómy, na ktoré sa hmyz prilepí, list sa zvinie a vylúči tráviace enzýmy.
- Mucholapka (Dionaea) chytá korisť aktívnym, mimoriadne rýchlym zaklapnutím dvoch polovíc listu.
- Krčiažnik (Nepenthes) láka hmyz do pasívnych pascí v tvare krčahov naplnených tráviacimi šťavami.
Mixotrofia je veľmi bežná aj u jednobunkových organizmov a rias.

Sekundárny metabolizmus rastlín
Sekundárne metabolity sú organické zlúčeniny produkované rastlinami, ktoré nie sú priamo nevyhnutné pre ich prežitie a rast v štandardných podmienkach. Zohrávajú však kľúčovú úlohu pri adaptácii rastlín na ich prostredie. Tieto zlúčeniny poskytujú rastlinám rôzne výhody, vrátane:
- Obrana proti herbivorom a patogénom: Mnohé sekundárne metabolity pôsobia ako toxíny, repelenty alebo antibiotiká, ktoré chránia rastliny pred škodcami, hmyzom a patogénnymi mikroorganizmami.
- Interakcie s prostredím: Sekundárne metabolity sa podieľajú na rôznych interakciách s prostredím, ako je prilákanie opeľovačov (napr. pigmenty v kvetoch, prchavé látky vo vôňach), súťaženie s inými rastlinami (alelopatia) alebo ochrana pred UV žiarením.
- Adaptácia na stresové podmienky: Niektoré sekundárne metabolity pomáhajú rastlinám vyrovnať sa s environmentálnym stresom, ako je sucho, zasolenie alebo vysoké teploty.
Primárne a sekundárne metabolity rastlín | Vysvetlené s príkladmi 🌿🔬
Triedy sekundárnych metabolitov
Sekundárne metabolity rastlín tvoria rozsiahlu a chemicky rozmanitú skupinu zlúčenín. Môžu byť klasifikované do troch hlavných tried:
Terpény a terpenoidy
Terpény sú najpočetnejšou skupinou sekundárnych metabolitov v rastlinách, odvodené od izoprénových jednotiek. Ich štruktúrna rozmanitosť je obrovská, od jednoduchých monoterpénov po komplexné triterpény a tetraterpény.
- Monoterpény (C10): Zodpovedné za éterické oleje, vône rastlín a niektoré repelenty proti hmyzu. Príklady zahŕňajú mentol, gáfor a limonén.
- Seskviterpény (C15): Často sa vyskytujú v rastlinných extraktoch a môžu mať rôzne biologické aktivity, vrátane antimikrobiálnych a protirakovinových účinkov.
- Diterpény (C20): Zahŕňajú rastové hormóny ako giberelíny a niektoré toxíny.
- Triterpény (C30): Predstavujú základnú štruktúru mnohých steroidov a saponínov, ktoré majú obranné funkcie.
- Tetraterpény (C40): Patria sem karotenoidy, dôležité pigmenty v fotosyntéze a antioxidanty.
- Polyterpény: Dlhé reťazce izoprénových jednotiek, ako je prírodný kaučuk.
Terpenoidy sú deriváty terpénov, ktoré boli chemicky modifikované (napr. oxidáciou alebo preskupením). Mnohé z nich majú významné farmakologické vlastnosti.
Fenolické zlúčeniny
Fenolické zlúčeniny obsahujú fenolový kruh (benzénový kruh s hydroxylovou skupinou). Sú syntetizované prostredníctvom šikimátovej a fenylpropanoidovej dráhy.
- Jednoduché fenoly: Patria sem fenolové kyseliny, ktoré sa často vyskytujú v rastlinných bunkových stenách.
- Flavonoidy: Veľká skupina zlúčenín zodpovedných za mnohé farby kvetov a plodov (žltá, červená, modrá). Majú silné antioxidačné vlastnosti a hrajú úlohu v signalizácii rastlín.
- Antokyany: Podskupina flavonoidov, ktoré dávajú rastlinám červené, fialové a modré sfarbenie.
- Lignín: Komplexný polymér, ktorý dodáva pevnosť bunkovým stenám a je kľúčovou zložkou dreva.
- Taníny: Polyfenoly s horkou chuťou, ktoré pôsobia ako obrana proti herbivorom a majú adstringentné vlastnosti.
Fenolické zlúčeniny sú známe svojimi antioxidačnými, protizápalovými a antimikrobiálnymi účinkami.
Alkaloidy
Alkaloidy sú dusíkaté organické zlúčeniny, ktoré sa často vyskytujú v rastlinách a majú silné fyziologické účinky na živočíchy. Mnohé z nich sú toxické a používajú sa v medicíne alebo ako rekreačné drogy.
- Kofeín: Stimulant, ktorý sa nachádza v káve, čaji a kakae.
- Nikotín: Obsiahnutý v tabaku, pôsobí ako insekticíd a stimulant.
- Morfin: Silný analgetikum získaný z ópia.
- Kvinín: Používa sa na liečbu malárie.
- Atropín: Alkaloid z belladony, používaný v oftalmológii a ako antidotum.
Biosyntéza alkaloidov je komplexná a často zahŕňa šikimátovú dráhu ako počiatočný krok.
Biosyntetické dráhy sekundárnych metabolitov
Výroba sekundárnych metabolitov v rastlinách je výsledkom zložitých biochemických dráh, ktoré často začínajú v primárnom metabolizme. Kľúčové dráhy zahŕňajú:
- Šikimátová dráha: Vedie k tvorbe aromatických aminokyselín (fenylalanín, tyrozín, tryptofán), ktoré sú prekurzormi fenolických zlúčenín a niektorých alkaloidov.
- Fenylpropanoidová dráha: Vychádza z fenylalanínu a vedie k širokej škále fenolických zlúčenín, vrátane flavonoidov, lignínu a niektorých alkaloidov.
- Mevalonátová (MVA) a metylerytritolfosfátová (MEP) dráha: Tieto dve dráhy sú zodpovedné za syntézu izoprénových jednotiek, ktoré sú stavebnými kameňmi terpénov.
Enzýmy hrajú rozhodujúcu úlohu pri katalýze týchto reakcií, pričom špecifické enzýmy riadia tvorbu konkrétnych sekundárnych metabolitov.
Funkcie sekundárnych metabolitov v rastlinách
Hoci sekundárne metabolity nie sú nevyhnutné pre základné životné funkcie, poskytujú rastlinám kľúčové výhody pre prežitie a reprodukciu:
Obrana:
- Proti herbivorom: Mnohé alkaloidy a terpény sú toxické alebo nechutné pre bylinožravce. Taníny môžu viazať proteíny v tráviacom trakte bylinožravcov, čím znižujú ich nutričnú hodnotu potravy.
- Proti patogénom: Fenolické zlúčeniny a niektoré terpény majú antimikrobiálne vlastnosti, ktoré chránia rastliny pred baktériami a hubami.
Interakcie s prostredím:
- Opeľovanie a šírenie semien: Flavonoidy a antokyany poskytujú farbu kvetom a plodom, čím priťahujú opeľovače a rozširovateľov semien. Prchavé terpény vo vôňach tiež zohrávajú úlohu pri prilákaní.
- Alelopatia: Niektoré rastliny vylučujú sekundárne metabolity do prostredia, ktoré inhibujú rast iných rastlín, čím znižujú konkurenciu o zdroje.
- Ochrana pred UV žiarením: Flavonoidy a iné fenolické zlúčeniny absorbujú škodlivé UV žiarenie, čím chránia rastlinné tkanivá.
Adaptácia na environmentálny stres:
- Sucho a zasolenie: Niektoré sekundárne metabolity môžu pomôcť regulovať osmotický potenciál rastliny alebo chrániť bunkové štruktúry pred poškodením v podmienkach stresu.

Výskyt a distribúcia sekundárnych metabolitov
Sekundárne metabolity nie sú rovnomerne distribuované v rastlinách. Ich prítomnosť a koncentrácia sa môžu líšiť v závislosti od druhu rastliny, tkaniva, vývojového štádia a environmentálnych podmienok.
- Tkanivová špecifickosť: Niektoré metabolity sa nachádzajú prevažne v určitých tkanivách, napríklad alkaloidy v listoch a koreňoch, silice v kvetoch a plodoch.
- Vývojová špecifickosť: Produkcia niektorých sekundárnych metabolitov sa môže meniť počas životného cyklu rastliny, napríklad produkcia farbív v plodoch počas dozrievania.
- Environmentálna indukcia: Stresové faktory, ako je napadnutie škodcami, infekcia patogénmi alebo environmentálny stres, môžu viesť k zvýšenej produkcii sekundárnych metabolitov ako obranná reakcia.
Výskum a využitie sekundárnych metabolitov
Štúdium sekundárneho metabolizmu rastlín má významné praktické aplikácie v rôznych odvetviach:
- Farmaceutický priemysel: Mnohé lieky sú odvodené alebo inšpirované rastlinnými sekundárnymi metabolitmi (napr. aspirín z vŕbovej kôry, morfín z maku).
- Potravinársky priemysel: Flavonoidy a antokyany sa používajú ako prírodné farbivá a antioxidanty.
- Poľnohospodárstvo: Pochopenie sekundárneho metabolizmu môže pomôcť pri vývoji prírodných pesticídov a zlepšení odolnosti rastlín voči škodcom a chorobám.
- Kozmetický priemysel: Rastlinné extrakty bohaté na sekundárne metabolity sa používajú v kozmetických výrobkoch pre ich antioxidačné a protizápalové vlastnosti.
tags: #primarny #a #sekundarny #metabolizmus #v #rastlinnach