Výskum sa zameriava na molekulárne mechanizmy kontrolujúce energetický metabolizmus buniek tukového tkaniva, kostrového svalu a pečene, ako aj na zabezpečenie ich vzájomnej komunikácie. Cieľom je zvýšiť metabolickú aktivitu týchto tkanív s cieľom zvýšiť energetický výdaj a zlepšiť metabolické zdravie človeka, vzhľadom na celosvetovú pandémiu obezity a s ňou spojené chronické ochorenia ako diabetes mellitus 2. typu. Kľúčové oblasti záujmu zahŕňajú výskum hnedého tukového tkaniva a alternatívnych mechanizmov tvorby tepla, ako aj objasnenie patogenézy nealkoholového stukovatenia pečene na úrovni jednotlivých typov a populácií buniek. Na štúdium bunkového metabolizmu sa využívajú bunkové línie, myšie modely, genetické a farmakologické nástroje, a tiež najmodernejšie techniky ako RNA sekvenovanie novej generácie, metabolomické analýzy a úprava genómu pomocou CRISPR-Cas9.
Základné metabolické procesy
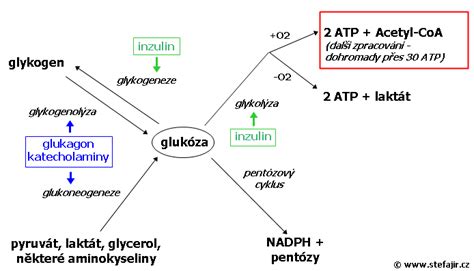
Glykolýza: Hlavná dráha metabolizmu glukózy
Glukóza patrí medzi základné zdroje energie. Glykolýza je kľúčový sled reakcií, ktorou sa v organizme katabolickou cestou z D-glukózy získava energia. Táto D-glukóza je pre niektoré bunky živinou, pre iné výhradným zdrojom energie. V jednoduchosti ide o premenu glukózy na pyruvát, ktorý po aktivácii vstupuje do Krebsovho cyklu.
Kroky glykolýzy
- V úvodnom kroku sa D-glukóza aktivuje fosforyláciou na glukóza-6-fosfát pomocou ATP.
- Tretím krokom je fosfofruktokinázová reakcia, ktorá je najvýznamnejšou a zároveň regulačnou reakciou celej glykolýzy. Dochádza k prenosu fosfátového zvyšku na fruktózu-6-fosfát v prítomnosti Mg²⁺ a ATP.
- Základným krokom je oxidácia glyceraldehydu-3-fosfátu v prítomnosti NAD⁺ za vzniku 1,3-bisfosfoglycerátu, ktorý je makroergickým medziproduktom. Z jednej molekuly fruktózy-1,6-bisfosfátu vznikajú dve molekuly glyceraldehydu-3-fosfátu, teda aj dve molekuly 1,3-bisfosfoglycerátu a 2× NADH + H⁺.
- V tejto reakcii vzniká po prvý raz energia vo forme ATP, keď enzým fosfoglycerátkináza prenesie fosfátový zvyšok na ADP za vzniku 3-fosfoglycerátu.
- V záverečnom kroku sa z fosfoenolpyruvátu prenáša posledný fosfátový zvyšok na ADP za vzniku ATP pomocou enzýmu pyruvátkináza.
V cykle 10 krokov vznikajú 2 molekuly pyruvátu, ktoré po aktivácii vstúpia do Krebsovho cyklu alebo sa môžu premieňať v cykloch anaeróbnej glykolýzy. Celkový energetický zisk (v podobe ATP) aeróbnej glykolýzy od glukózy po pyruvát je 2 ATP z 3. reakcie a 2 ATP zo 7. reakcie, spolu 4 ATP (čistý zisk 2 ATP, keďže 2 ATP boli spotrebované v prvých krokoch).
Glykogén: Zásobáreň glukózy
Glykogén je rozvetvený homopolymér molekúl glukózy, pričom väčšina glukózových zvyškov je viazaná pomocou α 1→4 väzieb. Každý dvanásty glukózový zvyšok je pripojený pomocou α 1→6 väzby, čo vytvára miesta rozvetvenia. Glykogén slúži u živočíchov ako zásobáreň sacharidov, z ktorej sa štiepením môžu uvoľňovať estery glukózy. Bohato hydratované glykogenové granuly sa nachádzajú v cytoplazme všetkých buniek. Ľudské telo môže skladovať približne 450 g glykogénu, z čoho 80-100 g je v pečeni (tzv. jaterný glykogén), ktorý sa využíva na udržiavanie stálej hladiny glukózy v krvi (glykémie). Ďalších 300 g je vo svalových bunkách (tzv. svalový glykogén), ktorý slúži ako interná svalová energetická zásoba pri svalovej práci. Svalové bunky neobsahujú glukóza-6-fosfatázu, preto nemôžu uvoľňovať čistú glukózu do krvného obehu.
Syntéza a degradácia glykogénu
Proces syntézy glykogénu prebieha v cytozole, intenzívny je hlavne v pečeni a kosternom svalstve. Vychádza z molekúl glukózy a vyžaduje primer. Tvorba glykosidových väzieb je endergonický proces, vyžadujúci energeticky bohaté substráty. Pomocou vetviaceho enzýmu (amylo-(1,4-1,6)-transglykosyláza) sa odštiepi oligosacharidový štěp, ktorý sa pripojí na C6 molekuly glukózy vo vnútri reťazca glykogénu, čím vzniká α 1→6 väzba. Syntéza prebieha v čase dostatočného prísunu energetických substrátov.
Hlavným regulačným enzýmom je glykogensyntáza, ktorej aktivita sa reguluje fosforyláciou - fosforylovaný enzým je neaktívny, defosforylácia vedie k aktivácii. Fosforyláciu ovplyvňuje pomer inzulín/glukagón.
Glykogén nie je nikdy úplne degradovaný. Degradácia prebieha v cytosole formou fosforolytického štiepenia pomocou enzýmu glykogenfosforyláza (fosforyláza) z neredukujúcich koncov. Uvoľňujú sa jednotlivé monomérne jednotky glukózy vo forme Glc-1-P (Coriho ester). Bohato rozvetvená molekula glykogénu sa odbouráva rýchlo.
Hlavným regulačným enzýmom glykogenolýzy je glykogenfosforyláza. Je aktívna vo fosforylovanej forme. Fosforyláciu katalyzuje fosforylánakináza, defosforyláciu proteínfosfatázy. Glykogenolýzu aktivujú kontraregulačné hormóny - glukagón, katecholamíny a glukokortikoidy. Vo svalových bunkách sa regulácia spája aj so zmenou koncentrácie Ca²⁺ iónov.
Vrodené poruchy metabolizmu glykogénu sa nazývajú glykogenózy, pri ktorých dochádza k hromadeniu glykogénu v bunkách, čo môže viesť k rôznym príznakom.
Glukoneogenéza: Syntéza glukózy z necukrových zdrojov
Glukoneogenéza (GNG) je metabolická dráha, ktorá vedie k vytvoreniu glukózy z niektorých nesacharidových uhlíkových substrátov. Tento proces prebieha vo všetkých živých organizmoch, u stavovcov hlavne v pečeni a v menšej miere v kôre obličiek. U ľudí môže substrát pre glukoneogenézu pochádzať z akýchkoľvek nesacharidových zdrojov, ktoré je možné premeniť na pyruvát alebo na niektorý z medziproduktov glykolýzy. Medzi hlavné prekurzory glukoneogenézy patria laktát, glycerol, alanín a glutamín.
Glukoneogenéza je silne endergonická, ale spojením s hydrolýzou ATP alebo GTP sa stáva exergonickou. Napríklad dráha od pyruvátu ku glukóza-6-fosfátu vyžaduje 4 molekuly ATP a 2 molekuly GTP.
Substráty a mechanizmy glukoneogenézy
Laktát je transportovaný do pečene v Coriho cykle, kde je pomocou laktátdehydrogenázy premenený na pyruvát. Transaminácia alebo deaminácia aminokyselín umožňuje vstup ich uhlíkovej kostry do cyklu.
Mastné kyseliny s nepárnym počtom uhlíkov sa môžu oxidovať na acetylkoenzým A a propionylkoenzým A, pričom propionylkoenzým A môže fungovať ako prekurzor sukcinylkoenzýmu A a následne vstúpiť do glukoneogenézy. Mastné kyseliny s párnym počtom uhlíkov sú oxidované len na acetylkoenzým A. U ľudí nebola existencia glyoxylátového cyklu potvrdená, preto sa predpokladá, že mastné kyseliny s párnym počtom uhlíkov nie je možné priamo premeniť na glukózu.
Počas ketózy sa acetylkoenzým A z mastných kyselín využíva na tvorbu ketolátok, ktoré môžu tvoriť až 11 % glukoneogenézy počas hladovania.
Regulácia glukoneogenézy
Glukoneogenéza je dráha pozostávajúca z 11 reakcií katalyzovaných enzýmami. Kľúčové kroky, ktoré sú v glykolýze nevratné (fosfofruktokinázová, pyruvátkinázová reakcia), sú v glukoneogenéze nahradené inými enzýmami (napr. fruktóza-1,6-bisfosfatáza, pyruvátkarboxyláza a PEP karboxykináza). Tieto enzýmy sú regulované podobnými molekulami ako glykolytické enzýmy, ale s opačnými výsledkami, čím sa zabraňuje zbytočným cyklom syntézy a rozkladu glukózy.
Globálna regulácia je sprostredkovaná glukagónom (ktorý sa vylučuje pri nízkej hladine glukózy v krvi), ktorý stimuluje glukoneogenézu. Inzulín pôsobí proti glukagónu inhibíciou glukoneogenézy. Metformín tiež potláča hepatálnu produkciu glukózy.
Metabolizmus glukózy v malígnych bunkách
Malígne epitelové bunky, podobne ako iné nádorové bunky, vykazujú zmenený metabolizmus glukózy, ktorý im umožňuje rýchlu proliferáciu a prežitie v náročnom mikroprostredí nádoru. Karcinogenéza je zložitý proces, ktorý zahŕňa mutácie v onkogénoch a tumor-supresorových génoch, a je ovplyvnený mikroprostredím nádoru.
Warburgov efekt a zvýšená spotreba glukózy
Viac ako 100 rokov po opísaní „Pasteurovho efektu“ (inhibícia oxidatívnej fosforylácie a prepnutie na glykolýzu v anaeróbnych podmienkach) zistil Otto Warburg, že nádorové bunky, na rozdiel od normálnych, využívajú glykolýzu namiesto mitochondriálnej oxidatívnej fosforylácie, a to aj v podmienkach s dostatkom kyslíka. Tento jav sa nazýva Warburgov efekt alebo aeróbna glykolýza.
Nádorové bunky sú nútené nadmerne vychytávať glukózu (GLU) na splnenie potrieb ich biosyntézy, zvýšenej energie a redoxnej potreby. Hoci produkcia ATP prostredníctvom glykolýzy je rýchlejšia, je menej účinná z hľadiska ATP vytvoreného na jednotku spotrebovanej GLU.
Energeticky nezávislý transport GLU cez plazmatickú membránu je sprostredkovaný glukózovými transportérmi (GLUT), ktoré sa líšia v tkanivovej distribúcii a afinite pre GLU. Tieto transportéry sú exprimované v membránach takmer všetkých typov buniek.

Hypoxia a regulácia metabolizmu
V dôsledku rýchlej proliferácie nádorových buniek dochádza v ich okolí k hypoxii, pretože lokálny cievny systém nie je schopný zabezpečiť dostatočné okysličenie. V týchto podmienkach musia nádorové bunky využívať vysoko regulované glykolytické cesty na produkciu ATP. K tomu dochádza prostredníctvom indukcie hypoxiou-indukovaného faktora-1 (HIF-1).
Produkcia laktátu a okyslenie mikroprostredia
Fyziologickým dôsledkom zvýšenej tvorby laktátu v nádoroch je pokles intersticiálneho pH. Kyslé prostredie sprostredkováva nádorovú invazivitu. Laktát stimuluje tvorbu kyseliny hyaluronovej a expresiu CD44, ktorý je hlavným transmembránovým receptorom. Naviazanie kyseliny hyalurónovej na CD44 na povrchu bunky znižuje bunkovú adherenciu, čo umožňuje samostatný pohyb buniek a zvyšuje malígnu progresiu.
Kyselina mliečna produkovaná nádorom pri glykolýze pôsobí ťažkosti T-lymfocytom - bunkám imunitného systému, a pôsobí ako antioxidant, čím znižuje účinky chemoterapie a rádioterapie.
Alternatívne zdroje energie: Glutamín
S rastom nádoru niektoré druhy rakoviny menia svoj hlavný zdroj potravy na glutamín, neesenciálnu aminokyselinu. Glutamín podporuje nárast hmotnosti u onkologických pacientov, ale pre rakovinu je nenahraditeľný a vedie k „kŕmeniu nádoru“.
Strategie liečby zamerané na metabolizmus
Liečba rakoviny zahŕňa blokovanie možných zdrojov výživy pre nádorové bunky a potláčanie metabolizmu glukózy a glutamínu. Potenciálne stratégie zahŕňajú:
- Nízkosacharidovú diétu s vynechaním potravín s vysokým obsahom glutamínu.
- Prírodné doplnky ako kvalitný zelený čaj (Matcha, Sencha), resveratrol, vitamín C, betaglukány.
- Zdravú, tzv. živú stravu s prevahou zásadotvorných potravín.
- Ovocné a zeleninové šťavy, pravidelné pôsty.
Silný imunitný systém dokáže bojovať s rakovinovými bunkami, ale niekedy potrebuje podporu. Dôležité je zablokovať všetky možné zdroje výživy pre nádorové bunky a potláčať metabolizmus glukózy a glutamínu vhodnými spôsobmi.
Nástroj na interpretáciu metabolizmu glukózy - precízna diagnostika
tags: #maligne #epitelialne #bunky #a #metabolizmus #glukozy